Как используется энергия в атф. Образование энергии в клетке. Систематическое наименование АТФ
text_fields
text_fields
arrow_upward
Извлечение энергии из питательных веществ - углеводов, белков, жиров происходит, в основном, внутри клетки. В ней все углеводы представлены глюкозой, белки - аминокислотами, жиры - жирными кислотами. В клетке глюкоза под влиянием энзимов цитоплазмы превращается в пировиноградную кислоту (в ходе анаэробного гликолиза) (рис. 1.6).
Рис. 1.6 Образование АТФ при полном окислении глюкозы
В ходе этих превращений из одной молекулы глюкозы образуется 2 молекулы АТФ (не считая 2 молекул АТФ, фосфорилирующих субстрат). Превращение пирувата в 2 молекулы ацетилкоэнзима А (АцКоА) способствует образованию еще 6 молекул АТФ. И, наконец, АцКоА поступает в митохондрии и, окисляясь в них до СО 2 и Н 2 О, образует еще 24 молекулы АТФ. Но не только пировиноградная кислота, а и жирные кислоты и большинство аминокислот превращаются в цитоплазме в АцКоА и также поступают в матрике митохондрий. В цикле Кребса АцКоА расщепляется до атомов водорода и окиси углерода. Окись углерода диффундирует из митохондрий, и далее из клетки. Атомы водорода соединяются с окисленным никотинамидадениндинуклеотидом (НАД+), формируя восстановленный НАД (НАДН), и с окисленным никотинамидаде-ниндинуклеотид фосфатом (НАДФ), формируя восстановленный НАДФН, а затем переносятся молекулами - переносчиками водорода от НАДН и НАДФН на систему ферментов внутренней митохондриальной мембраны.
В результате НАДН и НАДФН отдают один протон и два электрона в электротранспортную цепь, образуемую этими ферментами (рис.1.7).
 Рис.1.7 Взаимоотношения расщепления пищевых веществ и электрон-транспортной системы в клетке
Рис.1.7 Взаимоотношения расщепления пищевых веществ и электрон-транспортной системы в клетке
В ходе передачи электронов в цепи переносчиков возрастают окислительно-восстановительные потенциалы - от отрицательных значений до потенциала восстановления О 2 . Эта разница окислительно-восстановительных потенциалов и образует ту движущую силу, которая приводит к синтезу АТФ. Описанный перенос электронов и протонов от НАДН и НАДФН по цепи транспорта электронов называется окислительным фосфорилированием. Согласно хемиосмотической теории, объясняющей механизм образования энергии при окислительном фосфорилировании, в ходе передачи электронов по электронно-транспортной цепи, пара электронов три раза пересекает внутреннюю мембрану митохондрий, каждый раз перенося два протона наружу (рис. 1.8).
Рис. 1.8 Хемиосмотический механизм окислительного фосфорилирования во внутренней мембране митоходрий.В результате возникает высокая концентрация протонов снаружи мембраны, и низкая - в матриксе митохондрий и, как следствие, разница в электрическом потенциале между наружным (имеющим положительный заряд) и внутренним (накапливающим отрицательный заряд) слоем мембраны. Оба эти фактора (электрическое поле и разность концентраций) формируют электрохимический трансмембранный протонный градиент, благодаря которому протоны начинают возвращаться назад через мембрану. Это обратное движение протонов осуществляется через мембранный белок, к которому присоединяется АТФ-синтетаза, расположенная на внутренней (матричной) стороне мембраны. Взаимодействие мембранного белка с АТФ-синтетазой активирует ее и сопровождается синтезом АТФ из аденозин-дифосфорной (АДФ) и фосфорной кислот (Фн). Следовательно, поток протонов через мембрану активирует реакцию:
АДФ + Фн —> АТФ + Н 2 О
Энергия протонного градиента также обеспечивает транспорт ионов кальция и натрия через мембрану митохондрий, восстановление в них НАДФ+ с помощью НАДН, образование тепла. Молекулы АТФ, образовавшиеся в ходе гликолиза и окислительного фосфорилирования используются клеткой для обеспечения энергией почти всех внутриклеточных метаболических реакций.
Рис. 1.9 Схема молекулы АТФ. Стрелками показаны Тpuфocфam Высокоэнергетические связи.Макроэргические фосфатные связи молекулы АТФ очень нестойки и концевые фосфатные группы легко отщепляются от АТФ, освобождая энергию (7-10 ккал/моль АТФ) (рис. 1.9).
Энергия передается переносом отщепившихся, богатых энергией фосфатных групп на различные субстраты, ферменты, активируя их, расходуется на мышечное сокращение и т.п.
Энергетическая фосфогенная система
text_fields
text_fields
arrow_upward
Энергия макроэргических связей молекулы АТФ является универсальной формой запаса свободной энергии в организме. Вместе с тем, количество АТФ, хранимое внутри клетки невелико. Оно обеспечивает ее работу лишь в течение нескольких секунд. Это обстоятельство привело к формированию чувствительных механизмов, регулирующих энергетический обмен в скелетной, сердечной и нервных клетках. В этих тканях присутствуют органические фосфатные соединения, накапливающие энергию в форме фосфатных связей и представляющие собой источник этих богатых энергией фосфатных групп для синтеза АТФ. Органические фосфатные соединения получили название фосфагенов. Наиболее важным из них у человека является креатинфосфат (КФ). При его расщеплении высвобождается энергия до 10 ккал/моль, используемая для ресинтеза АТФ. Снижение содержания АТФ в этих тканях ведет к распаду КФ, а увеличение концентрации АТФ - к его ресинтезу. Так, в скелетной мышце концентрация КФ в 3-5 раз больше, чем АТФ. Гидролиз КФ (на креатин и фосфат) под влиянием фермента креатинкиназы обеспечивает ресинтез АТФ, являющейся источником энергии для мышечного сокращения:
Освободившийся креатин вновь используется клеткой для аккумуляции энергии в креатинфосфате. Этот эффект сохраняет концентрацию АТФ в клетке на относительно постоянном уровне. Поэтому фосфокреатин клеток скелетной мышцы и ее АТФ составляют, так называемую, энергетическую фосфогенную систему. Энергия фосфогенной системы используется для обеспечения «рывковой» мышечной активности, продолжительностью до 10-15 секунд, т.е. максимальной мышечной мощности, достаточной для бега на 100-метровую дистанцию.
Энергообеспечивающая система "гликоген-молочная кислота"
text_fields
text_fields
arrow_upward
Продолжающаяся более 10-15 секунд мышечная работа на максимально высоком уровне в следующие 30-40 секунд обеспечивается энергией анаэробного гликолиза, т.е. превращением молекулы глюкозы из расщепляющегося углеводного депо - гликогена печени и мышц до молочной кислоты. При анаэробном гликолизе молекулы АТФ образуются почти в 2,5 раза быстрее, чем при аэробном окислении в митохондриях. Таким образом, фосфогенная система и анаэробное расщепление гликогена до молочной кислоты (система гликоген - молочная кислота) обеспечивают человеку возможность мышечной рывковой работы значительного объема (в спорте - бег на короткие дистанции, подъем тяжестей, ныряние и т.д.) Более продолжительная мышечная работа человека требует усиления окислительного фосфорилирования в митохондриях, обеспечивающего, как было показано выше, основную часть ресинтеза АТФ.
Как именно энергия запасается в АТФ (аденозинтрифосфат), и как она отдается для совершения какой-то полезной работы? Кажется невероятно сложным, что некая абстрактная энергия вдруг получает материальный носитель в виде молекулы, находящейся внутри живых клеток, и что она может высвобождаться не в виде тепла (что более-менее понятно), а в виде создания другой молекулы. Обычно авторы учебников ограничиваются фразой «энергия запасается в виде высокоэнергетической связи между частями молекулы, и отдается при разрыве этой связи, совершая полезную работу», но это ничего не объясняет.

В самых общих чертах эти манипуляции с молекулами и энергией происходят так: сначала . Или создаются в хлоропластах в цепи похожих реакций. На это тратится энергия, получаемая при контролируемом сгорании питательных веществ прямо внутри митохондрий или энергия фотонов солнечного света, падающих на молекулу хлорофилла. Потом АТФ доставляется в те места клетки, где необходимо совершить какую-то работу. И при отщеплении от нее одной или двух фосфатных групп выделяется энергия, которая эту работу и совершает. АТФ при этом распадается на две молекулы: если отщепилась только одна фосфатная группа, то АТФ превращается в АДФ (аденозинДИфосфат, отличающийся от аденозинТРИфосфата только отсутствием той самой отделившейся фосфатной группы). Если АТФ отдала сразу две фосфатные группы, то энергии выделяется больше, а от АТФ остается аденозинМОНОфосфат (АМФ ).
Очевидно, что клетке необходимо осуществлять и обратный процесс, превращая молекулы АДФ или АМФ в АТФ, чтобы цикл мог повториться. Но эти молекулы-«заготовки» могут спокойно плавать рядом с недостающими им для превращения в АТФ фосфатами, и никогда с ними не объединиться, потому что такая реакция объединения энергетически невыгодна.

Что такое «энергетическая выгода» химической реакции, понять довольно просто, если знать о втором законе термодинамики : во Вселенной или в любой системе, изолированной от остальных, беспорядок может лишь нарастать. То есть сложноорганизованные молекулы, сидящие в клетке в чинном порядке, в соответствии с этим законом могут только разрушаться, образуя более мелкие молекулы или даже распадаясь на отдельные атомы, ведь тогда порядка будет заметно меньше. Чтобы понять эту мысль, можно сравнить сложную молекулу с собранным из Лего самолетиком. Тогда мелкие молекулы, на которые распадается сложная, будут ассоциироваться с отдельными частями этого самолета, а атомы — с отдельными кубиками Лего. Посмотрев на аккуратно собранный самолет и сравнив его с беспорядочной кучей деталей, становится понятно, почему сложные молекулы содержат больше порядка, чем мелкие.
Такая реакция распада (молекул, не самолета) будет энергетически выгодной, а значит может осуществляться самопроизвольно, и при распаде будет выделяться энергия. Хотя на самом деле и расщепление самолета будет энергетически выгодно: несмотря на то, что сами по себе детали отщепляться друг от друга не будут и над их отцеплением придется попыхтеть сторонней силе в виде пацана, который хочет использовать эти детали для чего-то другого, он затратит на превращение самолета в хаотическую кучу деталей энергию, полученную от поедания высокоупорядоченной пищи. И чем плотнее слиплись детали, тем больше энергии будет потрачено, в том числе выделено в виде тепла. Итог: кусок плюшки (источник энергии) и самолет превращены в беспорядочную массу, молекулы воздуха вокруг ребенка нагрелись (а значит движутся более беспорядочно) — хаоса стало больше, то есть расщепление самолета энергетически выгодно.
Подводя итог, можно сформулировать такие правила, следующие из второго закона термодинамики:
1. При снижении количества порядка энергия выделяется, происходят энергетически выгодные реакции
2. При увеличении количества порядка энергия поглощается, происходят энергетически затратные реакции
На первый взгляд, такое неизбежное движение от порядка к хаосу делает невозможным обратные процессы, такие как построение из одной оплодотворенной яйцеклетки и молекул питательных веществ, поглощеных матерью-коровой, несомненно весьма упорядоченного по сравнению с пережеванной травой теленка.
Но все-таки это происходит, и причина этого в том, что живые организмы имеют одну фишку, позволяющую и поддержать стремление Вселенной к энтропии, и построить себя и свое потомство: они объединяют в один процесс две реакции, одна из которых энергетически выгодна, а другая энергозатратна . Таким совмещением двух реакций можно добиться того, чтобы энергия, выделяемая при первой реакции, с избытком перекрывала энергетические затраты второй. В примере с самолетом отдельно взятое его разбирание энергозатратно, и без стороннего источника энергии в виде разрушенной метаболизмом пацана плюшки самолет стоял бы вечно.

Это как при катании с горки на санках: сначала человек во время поглощения пищи запасает энергию, полученную в результате энергетически выгодных процессов расщепления высокоупорядоченной курицы на молекулы и атомы в его организме. А потом тратит эту энергию, затаскивая санки на гору. Перемещение санок от подножия к вершине энергетически невыгодно, поэтому самопроизвольно они туда никогда не закатятся, на это нужна какая-то сторонняя энергия. И если энергии, полученной от поедания курицы, будет недостаточно для преодоления подъема, то процесса «скатывание на санках с вершины горы» не будет.
Именно энергозатратные реакции (energy-consuming reaction ) увеличивают количество порядка, поглощая энергию, выделяемую при сопряженной реакции. И баланс между выделением и потреблением энергии в этих сопряженных реакциях всегда должен быть положительным, то есть их совокупность будет увеличивать количество хаоса. Примером увеличения энтропии (неупорядоченности) (entropy [‘entrə pɪ ] ) является выделение тепла при энергодающей реакции (energy supply reaction ): соседние с вступившими в реакцию молекулами частицы вещества получают энергичные толчки от реагирующих, начинают двигаться быстрее и хаотичнее, распихивая в свою очередь другие молекулы и атомы этого и соседних веществ.

Вернемся еще раз к получению энергии из пищи: кусок Banoffee Pie гораздо более упорядочен, чем получившаяся в результате пережевывания масса, попавшая в желудок. Которая в свою очередь состоит из крупных, более упорядоченных молекул, чем те, на которые ее расщепит кишечник. А они в свою очередь будут доставлены в клетки тела, где от них будут отрывать уже отдельные атомы и даже электроны… И на каждом этапе увеличения хаоса в отдельно взятом куске торта будет происходить выделение энергии, которую улавливают органы и органеллы счастливого поедателя, запасая ее в виде АТФ (энергозатратно), пуская на построение новых нужных молекул (энергозатратно) или на нагревание тела (тоже энергозатратно). В системе «человек — Banoffee Pie — Вселенная» порядка в результате этого стало меньше (за счет разрушения кейка и выделения тепловой энергии перерабатывающими его органеллами), но в отдельно взятом человеческом теле счастья порядка стало больше (за счет возникновения новых молекул, частей органелл и целых клеточных органов).
Если вернуться к молекуле АТФ, после всего этого термодинамического отступления становится понятно, что на создание ее из составных частей (более мелких молекул) необходимо затратить энергию, полученную от энергетически выгодных реакций. Один из способов ее создания подробно описан , другой (весьма схожий) используется в хлоропластах, где вместо энергии протонного градиента используется энергия фотонов, испущенных Солнцем.

Можно выделить три группы реакций, в результате которых производится АТФ (смотри схему справа):
- расщепление глюкозы и жирных кислот на крупные молекулы в цитоплазме уже позволяет получить некоторое количество АТФ (небольшое, на одну расщепленную на этом этапе молекулу глюкозы приходится всего лишь 2 полученные молекулы АТФ). Но основная цель этого этапа заключается в создании молекул, использующихся в дыхательной цепи митохондрий.
- дальнейшее расщепление полученных на предыдущем этапе молекул в цикле Кребса, протекающее в матриксе митохондрий, дает всего одну молекулу АТФ, его основная цель та же, что и в прошлом пункте.
- наконец накопленные на предыдущих этапах молекулы используются в дыхательной цепи митохондрий для производства АТФ, и вот тут его выделяется много (про это подробнее ниже).
Если описать все это более развернуто, взглянув на те же реакции с точки зрения получения и затрат энергии, получится вот что:
0. Молекулы пищи аккуратно сжигаются (окисляются) в первичном расщеплении, происходящем в цитоплазме клетки, а также в цепи химических реакций под названием «цикл Кребса», протекающем уже в матриксе митохондрий — энергодающая часть подготовительного этапа.
В результате сопряжения с этими энергетически выгодными реакциями других, уже энергетически невыгодных реакций создания новых молекул образуются 2 молекулы АТФ и несколько молекул других веществ — энергозатратная часть подготовительного этапа. Эти попутно образующиеся молекулы являются переносчиками высокоэнергетических электронов, которые будут использованы в дыхательной цепи митохондрий на следующем этапе.

1. На мембранах митохондрий, бактерий и некоторых архей происходит энергодающее отщепление протонов и электронов от молекул, полученных в предыдущем этапе (но не от АТФ). Прохождение электронов по комплексам дыхательной цепи (I, III и IV на схеме слева) показано желтыми извилистыми стрелками, прохождение через эти комплексы (а значит, и через внутреннюю мембрану митохондрии) протонов — красными стрелками.
Почему электроны нельзя просто отщепить от молекулы-переносчика с использованием мощного окислителя-кислорода и использовать выделяющуюся энергию? Зачем передавать их от одного комплекса к другому, ведь в итоге они к тому же кислороду и приходят? Оказывается, чем больше разница в способности притягивать электроны у электронодающей (восстановителя ) и электроноберущей (окислителя ) молекул, участвующих в реакции передачи электрона, тем большая энергия выделяется при этой реакции.
Разница в такой способности у образующихся в цикле Кребса молекул-переносчиков электронов и кислорода такова, что выделившейся при этом энергии было бы достаточно для синтеза нескольких молекул АТФ. Но из-за такого резкого перепада в энергии системы эта реакция протекала бы с почти взрывной мощью, и почти вся энергия выделялась бы в виде неулавливаемого тепла, то есть фактически терялась.
Живые клетки же делят эту реакцию на несколько маленьких стадий, сначала передавая электроны от слабо притягивающих молекул-носителей к чуть сильнее притягивающему первому комплексу в дыхательной цепи, от него к еще немного сильнее притягивающему убихинону (или коэнзиму Q-10 ), чья задача заключается в перетаскивании электронов к следующему, еще немного сильнее притягивающему дыхательному комплексу, который получает свою часть энергии от этого несостоявшегося взрыва, пуская ее на прокачку протонов через мембрану.. И так до момента, пока электроны не встретятся наконец с кислородом, притянувшись к нему, прихватив пару протонов, и не образуют молекулу воды. Такое деление одной мощной реакции на мелкие шаги позволяет почти половину полезной энергии направить на совершение полезной работы: в данном случае на создание протонного электрохимического градиента , о котором речь пойдет во втором пункте.
Как именно энергия передаваемых электронов помогает сопряженной энергозатратной реакции прокачки протонов через мембрану, сейчас только начинают выяснять. Скорее всего, присутствие электрически заряженной частицы (электрона) влияет на конфигурацию того места во встроенном в мембрану протеине, где он находится: так, что это изменение провоцирует затягивание протона в протеин и его движение через протеиновый канал в мембране. Важно то, что фактически энергия, полученная в результате отщепления высокоэнергетичных электронов от молекулы-носителя и итоговой передачи их кислороду, запасается в виде протонного градиента.
2. Энергия протонов, накопившихся в результате событий из пункта 1 с внешней стороны мембраны и стремящихся попасть на внутреннюю сторону, состоит из двух однонаправленных сил:
- электрической (положительный заряд протонов стремится перейти в место скопления отрицательных зарядов с другой стороны мембраны) и
- химической (как в случае любых других веществ, протоны пытаются равномерно рассеяться в пространстве, распространившись из мест с их высокой концентрацией в места, где их мало)

Электрическое притяжение протонов к отрицательно заряженной стороне внутренней мембраны является намного более мощной силой, чем возникающее из-за разницы в концентрации протонов их стремление перейти в место с меньшей концентрацией (это обозначено шириной стрелок на схеме вверху). Совместная энергия этих влекущих сил настолько велика, что ее хватает и на перемещение протонов внутрь мембраны, и на подпитывание сопутствующей энергозатратной реакции: создание АТФ из АДФ и фосфата.
Рассмотрим подробнее, почему на это нужна энергия, и как именно энергия стремления протонов превращается в энергию химической связи между двумя частями молекулы АТФ.
Молекула АДФ (на схеме справа) не жаждет обзаводиться еще одной фосфатной группой: тот атом кислорода, к которому эта группа может прикрепиться, заряжен так же отрицательно, как и фосфат, а значит они взаимно отталкиваются. И вообще АДФ не собирается вступать в реакции, она химически пассивна. У фосфата, в свою очередь, к тому атому фосфора, который мог бы стать местом связи фосфата и АДФ при создании молекулы АТФ, присоединен собственный атом кислорода, так что и он инициативы проявить не может.
Поэтому эти молекулы необходимо связать одним ферментом, развернуть их так, чтобы связи между ними и «лишними» атомами ослабли и разорвались, а после этого подвести два химически активных конца этих молекул, на которых атомы испытывают недостаток и избыток электронов, друг к другу.
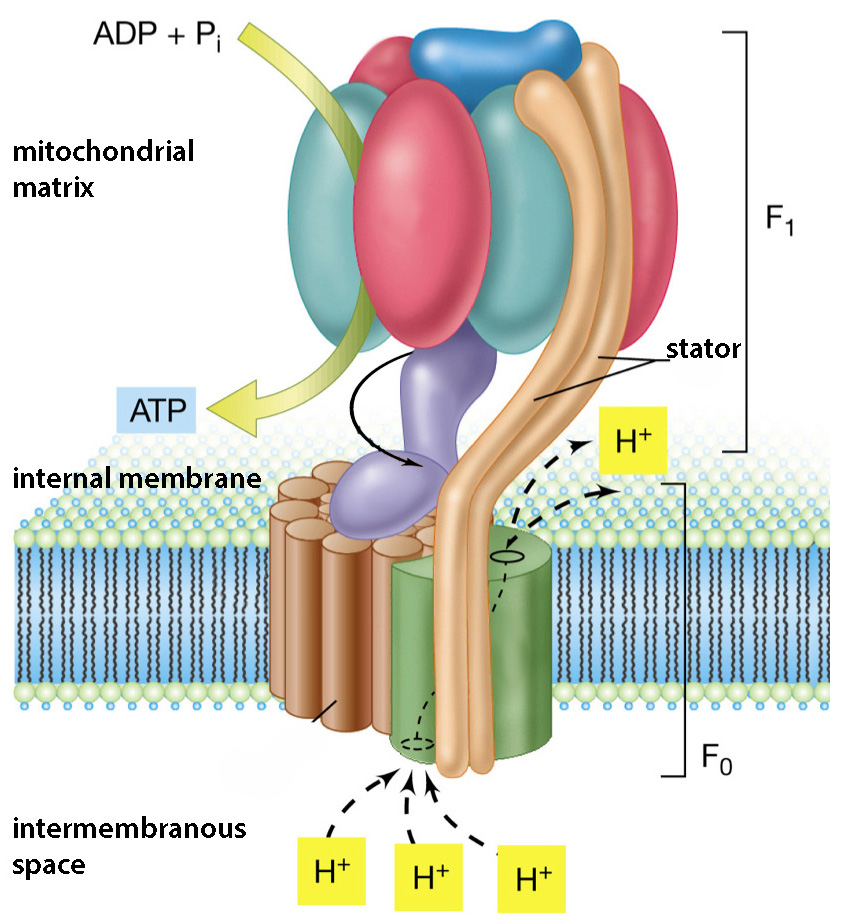
Попавшие в поле взаимной досягаемости ионы фосфора (P +) и кислорода (O —) связываются прочной ковалентной связью за счет того, что совместно овладевают одним электроном, изначально принадлежавшим кислороду. Этим обрабатывающим молекулы ферментом является АТФ-синтаза , а энергию на изменение и своей конфигурации, и взаимного расположения АДФ и фосфата она получает от проходящих через нее протонов. Протонам энергетически выгодно попасть на противоположно заряженную сторону мембраны, где к тому же их мало, а единственный путь проходит через фермент, «ротор» которого протоны попутно вращают.
Строение АТФ-синтазы показано на схеме справа. Ее вращающийся за счет прохождения протонов элемент выделен фиолетовым цветом, а на подвижной картинке внизу показана схема его вращения и создания при этом молекул АТФ. Фермент работает практически как молекулярный мотор, превращая электрохимическую энергию тока протонов в механическую энергию трения двух наборов протеинов друг о друга: вращающаяся «ножка» трется о неподвижные протеины «шляпки гриба», при этом субъединицы «шляпки» изменяют свою форму. Эта механическая деформация превращается в энергию химических связей при синтезе АТФ, когда молекулы АДФ и фосфата обрабатываются и разворачиваются нужным для образования между ними ковалентной связи образом.

Каждая АТФ-синтаза способна синтезировать до 100 молекул АТФ в секунду, и на каждую синтезируемую молекулу АТФ через синтетазу должно пройти около трех протонов. Большая часть синтезируемых в клетках АТФ образуется именно этим путем, и лишь небольшая часть является результатом первичной обработки молекул пищи, происходящей вне митохондрий.
В любой момент в типичной живой клетке находится примерно миллиард молекул АТФ. Во многих клетках вся эта АТФ сменяется (т.е. используется и создается вновь) каждые 1-2 минуты. Средний человек в состоянии покоя использует каждые 24 часа массу АТФ, примерно равную его собственной массе.

В целом почти половина энергии, выделяющаяся при окислении глюкозы или жирных кислот до углекислого газа и воды, улавливается и используется для протекания энергетически невыгодной реакции образования АТФ из АДФ и фосфатов. Коэффициент полезного действия в размере 50% — это очень неплохо, например двигатель автомобиля пускает на полезную работу всего лишь 20% содержащейся в топливе энергии. При этом остальная энергия в обоих случаях рассеивается в виде тепла, и так же как некоторые автомобили, животные постоянно тратят этот избыток (хоть и не полностью, конечно) на разогревание тела. В процессе упомянутых здесь реакций одна молекула глюкозы, постепенно расщепленная до углекислого газа и воды, поставляет клетке 30 молекул АТФ.
Итак, с тем, откуда берется энергия и как именно она запасается в АТФ, все более-менее понятно. Осталось понять, как именно запасенная энергия отдается и что при этом происходит на молекулярно-атомном уровне.

Образованная ковалентная связь между АДФ и фосфатом называется высокоэнергетичной по двум причинам:
- при ее разрушении выделяется много энергии
- электроны, участвующие в создании этой связи (то есть вращающиеся вокруг атомов кислорода и фосфора, между которыми эта связь образована) высокоэнергетичны, то есть находятся на «высоких» орбитах вокруг ядер атомов. И им было бы энергетически выгодно перескочить на уровень пониже, выделив излишек энергии, но пока они находятся именно в этом месте, скрепляя атомы кислорода и фосфора, «спрыгнуть» не получится.
Это стремление электронов упасть на более удобную низкоэнергетичную орбиту обеспечивает и легкость разрушения высокоэнергетичной связи, и выделяемую при этом в виде фотона (являющегося переносчиком электромагнитного взаимодействия) энергию. В зависимости от того, какие молекулы будут подставлены ферментами к разрушающейся молекуле АТФ, какая именно молекула поглотит испущенный электроном фотон, могут происходить разные варианты событий. Но каждый раз энергия, запасенная в виде высокоэнергетической связи, будет использоваться на какие-то нужды клетки :
Сценарий 1: фосфат может быть перенесен на молекулу другого вещества. При этом высокоэнергетичные электроны образуют новую связь, уже между фосфатом и крайним атомом этой молекулы-реципиента. Условием протекания такой реакции является ее энергетическая выгода: в этой новой связи электрон должен обладать немного меньшей энергией, чем когда он был частью молекулы АТФ, испустив часть энергии в виде фотона вовне.

Цель такой реакции заключается в активации молекулы-рецепиента (на схеме слева она обозначена В -ОН): до присоединения фосфата она была пассивной и не могла вступить в реакцию с другой пассивной молекулой А , но теперь она является обладателем запаса энергии в виде высокоэнергетичного электрона, а значит может ее куда-то потратить. Например, на то, чтобы присоединить к себе молекулу А , которую без такого финта ушами (то есть высокой энергии связующего электрона) присоединить невозможно. Фосфат при этом отсоединяется, сделав свое дело.
Получается такая цепочка реакций:
1. АТФ + пассивная молекула В ➡️ АДФ + активная за счет присоединенного фосфата молекула В-Р
2. активированная молекула В-Р + пассивная молекула А ➡️соединенные молекулы А-В + отщепившийся фосфат (Р )
Обе этих реакции энергетически выгодные: в каждой из них участвует высокоэнергетичный связующий электрон, который при разрушении одной связи и построении другой теряет часть своей энергии в виде испускания фотона. В результате этих реакций соединились две пассивные молекулы. Если рассмотреть реакцию соединения этих молекул напрямую (пассивная молекула В + пассивная молекула А ➡️соединенные молекулы А-В ), то она оказывается энергетически затратной, и совершиться не может. Клетки «совершают невозможное», сопрягая эту реакцию с энергетически выгодной реакцией расщепления АТФ на АДФ и фосфат во время совершения тех двух реакций, которые описаны выше. Отщепление происходит в два этапа, на каждом из которых часть энергии связующего электрона тратится на совершение полезной работы, а именно на создание нужных связей между двумя молекулами, из которых получается третья (А-В ), необходимая для функционирования клетки.
Сценарий 2: фосфат может быть отщеплен одномоментно от молекулы АТФ, а выделяющаяся энергия улавливается ферментом или рабочим протеином и тратится на совершение полезной работы.

Как можно уловить что-то настолько неощутимое, как ничтожное возмущение электромагнитного поля в момент падения электрона на более низкую орбиту? Очень просто: с помощью других электронов и с помощью атомов, способных поглотить выделяемый при этом электроном фотон.

Атомы, составляющие молекулы, скреплены в прочные цепочки и кольца за счет (такую цепочку представляет собой несвернутый протеин на картинке справа). А отдельные части этих молекул притянуты друг к другу более слабыми электромагнитными взаимодействиями (например, водородными связями или силами Ван дер Ваальса), что и позволяет им сфорачиваться в сложные структуры. Некоторые из этих конфигураций атомов очень стабильны, и никакое возмущение электромагнитного поля их не поколебит.. не поколебёт.. в общем, они устойчивы. А некоторые довольно подвижны, и достаточно легкого электромагнитного пинка, чтобы они изменили свою конфигурацию (обычно это не ковалентные связи). И именно такой пинок дает им тот самый прилетевший фотон-переносчик электромагнитного поля, испущенный перешедшим на более низкую орбиту электроном при отсоединении фосфата.
Изменения конфигурации протеинов в результате расщепления молекул АТФ ответственны за самые удивительные события, происходящие в клетке. Наверняка те, кто интересуются клеточными процессами хотя бы на уровне «посмотрю их анимацию на youtube» натыкались на видео, показывающее протеиновую молекулу кинезина , в прямом смысле слова шагающую, переставляя ноги, по нити клеточного скелета, перетаскивая присоединенный к ней груз.
Именно отщепление фосфата от АТФ обеспечивает это шагание, и вот каким образом:

Кинезин (kinesin ) относится к особому виду протеинов, которым свойственно спонтанно менять свою конформацию (взаимное положение атомов в молекуле). Оставленный в покое, он случайным образом переходит из конформации 1, в которой он прикреплен одной «ногой» к актиновому филаменту (actin filament ) — самой тонкой нити, образующей цитоскелет клетки (cytoskeleton ), в конформацию 2, сделав таким образом шаг вперед и стоя на двух «ногах». Из конформации 2 он с равной вероятностью перейдет как в конформацию 3 (приставляет заднюю ногу к передней), так и обратно в конформацию 1. Поэтому движения кинезина в каком-либо направлении не происходит, он просто бесцельно фланирует.

Но все меняется, стоит ему соединиться с молекулой АТФ. Как показано на схеме слева, присоединение АТФ к кинезину, находящемуся в конформации 1, приводит к изменению его пространственного положения и он переходит в конформацию 2. Причина этого — взаимное электромагнитное влияние молекул АТФ и кинезина друг на друга. Эта реакция является обратимой, потому что энергии затрачено не было, и если АТФ отсоединится от кинезина, он просто поднимет «ногу», оставшись на месте, и будет ждать следующую молекулу АТФ.
Но если она задержится, то из-за взаимного притяжения этих молекул связь, удерживающая фосфат в пределах АТФ, разрушается. Выделившаяся при этом энергия, а так же распад АТФ на две молекулы (которые уже по другому влияют своими электромагнитными полями на атомы кинезина) приводят к тому, что конформация кинезина меняется: он «подтаскивает заднюю ногу». Осталось сделать шаг вперед, что и происходит при отсоединении АДФ и фосфата, возвращающем кинезин в исходную конформацию 1.
В результате гидролиза АТФ кинезин сдвинулся вправо, и как только к нему присоединится следующая молекула, он сделает еще одну пару шагов, использовав запасенную в ней энергию.
Важно, что кинезин, находящийся в конформации 3 с присоединенными АДФ и фосфатом не может вернуться в конформацию 2, сделав «шаг назад». Это объясняется все тем же принципом соответствия второму закону терморегуляции: переход системы «кинезин + АТФ» из конформации 2 в конформацию 3 сопровождается выделением энергии, а значит обратный переход будет энергозатратным. Чтобы он произошел, нужно откуда-то взять энергию на соединение АДФ с фосфатом, а взять ее в этой ситуации неоткуда. Поэтому соединенному с АТФ кинезину открыт путь только в одну сторону, что и позволяет совершать полезную работу по перетаскиванию чего-либо из одного конца клетки в другой. Кинезин например участвует в растаскивании хромосом делящейся клетки при митозе (процессе деления эукариотических клеток). А мышечный протеин миозин бежит вдоль актиновых филаментов, вызывая сокращение мышцы.

Это движение бывает очень быстрым: некоторые моторные (отвечающие за различные формы клеточной подвижности) протеины, задействованные в репликации генов, мчатся вдоль цепочки ДНК со скоростью тысячи нуклеотидов в секунду.
Все они передвигаются за счет гидролиза АТФ(разрушения молекулы с присоединением к получающимся в результате распада меньшим молекулам атомов, взятых из молекулы воды. Гидролиз показан на правой части схемы взаимопревращения АТФ и АДФ). Или за счет гидролиза ГТФ , отличающегося от АТФ только тем, что в его состав входит другой нуклеотид (гуанин).
Сценарий 3 : отщепление от АТФ или другой подобной молекулы, содержащей нуклеотид, сразу двух фосфатных групп приводит к еще большему выбросу энергии, чем когда отщепляется только один фосфат. Такой мощный выброс позволяет создавать прочный сахарофосфатный остов молекул ДНК и РНК:

1. для того, чтобы нуклеотиды могли присоединяться к строящейся цепи ДНК или РНК, их нужно активировать, присоединив две молекулы фосфата. Это энергозатратная реакция, выполняемая клеточными ферментами.
2. фермент ДНК- или РНК-полимераза (на схеме внизу не показан) присоединяет активированный нуклеотид (на схеме показан ГТФ) к строящемуся полинуклеотиду и катализирует отщепление двух фосфатных групп. Выделившаяся энергия используется на создание связи между фосфатной группой одного нуклеотида и рибозой другого. Созданные в результате связи не являются высокоэнергетичными, а значит разрушить их не просто, что является преимуществом для построения молекулы, содержащей наследственную информацию клетки или передающей ее.

В природе возможно спонтанное протекание только энергетически выгодных реакций, что обусловлено вторым законом термодинамики
Тем не менее живые клетки могут совмещать две реакции, одна из которых дает чуть больше энергии, чем поглощает вторая, и таким образом осуществлять энергозатратные реакции. Энергозатратные реакции направлены на создание из отдельных молекул и атомов более крупных молекул, клеточных органелл и целых клеток, тканей, органов и многоклеточных живых существ, а так же на запасание энергии для их метаболизма
Запасание энергии осуществляется за счет контролируемого и постепенного разрушения органических молекул (энергодающий процесс), сопряженного с созданием молекул-энергоносителей (энергозатратный процесс). Фотосинтезирующие организмы запасают таким образом энергию улавливаемых хлорофиллом солнечных фотонов
Молекулы-энергоносители делятся на две группы: хранящие энергию в виде высокоэнергетической связи или в виде присоединенного высокоэнергетического электрона. Впрочем, в первой группе высокая энергия обеспечивается таким же высокоэнергетическим электроном, так что можно сказать, что энергия запасается в загнанных на высокий уровень электронах, находящихся в составе разных молекул
Запасенная таким образом энергия отдается так же двумя способами: разрушением высокоэнергетической связи или передачей высокоэнергетических электронов для постепенного снижения их энергии. В обоих случаях энергия выделяется в виде испускания переходящим на более низкий энергетический уровень электроном частицы-переносчика электромагнитного поля (фотона) и тепла. Этот фотон улавливается таким образом, чтобы была совершена полезная работа (образование нужной для метаболизма молекулы в первом случае и прокачки протонов через мембрану митохондрии во втором)
Запасенная в виде протонного градиента энергия используется для синтеза АТФ, а также для других клеточных процессов, которые остались за рамками этой главы (думаю, никто не в обиде, учитывая ее размер). А синтезированная АТФ используется так, как описано в предыдущем пункте.
Поступающая в организм человека пища претерпевает сложные химические превращения, т.е. частично подвергается окислению или анаэробному распаду. При анаэробном распаде освобождается химическая энергия, необходимая для движения, а также для синтеза необходимых для организма веществ.
Обмен веществ (метаболизм) в живых организмах состоит из двух связанных между собой процессов:
- анаболизма
- катаболизма
Анаболизм или ассимиляция – синтез из простых более сложных соединений на основе поступающих в организм из внешней среды веществ.
Например, органические вещества в зеленых растениях образуются в результате фотосинтеза из углекислого газа и воды.
Катаболизм или диссимиляция – процесс, обратный анаболизму. При катаболизме происходит разложение сложных соединений на более простые, которые затем выделяются как конечные продукты в окружающую среду.
При катаболизме основным источником углеводов являются углеводы, которые расщепляются гидролитическими ферментами. Если у растений при прорастании семян крахмал подвергается гидролизу ферментом амилазой, с образованием дисахарида мальтозы, то у животных под действием амилазы слюны и поджелудочной железы, образуя мальтозу. Далее мальтоза под действием фермента мальтазы переходит в глюкозу, которая в результате брожения, гликолиза и дыхания в конечном итоге расщепляется до углекислоты и воды. Энергия, выделяемая при этих процессах, аккумулируется в организме. Установлено, что при сгорании одного грамма углеводов выделяется 4,1 ккал (17,22 кДж).
Катаболизм жиров и белков также начинается с их гидролитического расщепления под влиянием специфических ферментов, с образованием в первом случае свободных жирных кислот и глицерина, во втором – низкомолекулярных пептидов и аминокислот.
Метаболизм или обмен веществ можно разделить на три этапа:
- Первый- это пищеварение, который заключается в механической и химической обработке пищи в пищеварительных органах и всасывание питательных веществ.
- Второй этап это – промежуточный обмен, который включает процессы распада и синтеза веществ. Этот процесс сопровождается образованием промежуточных и конечных продуктов обмена. Например, глюкоза прежде чем превратиться в конечные продукты обмена СО2 и Н2О, претерпевает ряд промежуточных превращений.
- Третий этап – выделение продуктов метаболизма из организма с выдыхаемым воздухом, мочой и т.д. Вещества, влияющие на течение реакции обмена веществ называют метаболитами. К ним относятся аминокислоты, жирные кислоты, сахара, азотистые основания и другие соединения.
Метаболизм или обмен веществ неразрывно связан с превращением энергии. Живой организм постоянно нуждается в поступлении энергии из внешней среды. Было установлено, что при фотосинтезе, т.е. преобразовании энергии солнечного света, последняя запасается в виде потенциальной химической энергии в органических веществах. Потенциальная химическая энергия, которая образуется в результате распада углеводов, жиров и других высокомолекулярных соединений накапливается или аккумулируется в макроэргических соединениях.
В процессах обмена энергия выделяется следующим образом. Вначале высокомолекулярные вещества гидролитически распадаются на низкомолекулярные; например, полисахариды – до моносахаридов; белки – до аминокислот; жиры – до жирных кислот и глицерина. При этом энергия, выделяющаяся при гидролитическом распаде этих веществ очень незначительна. Далее происходит выделение большого количества энергии в процессе гликолиза, окисления жирных кислот, аминокислот. Из продуктов гидролиза основное энергетическое значение имеют три: ацетилкоэнзим А, В -кетоглутаровая кислота и щавелево-уксусная кислота. Эти вещества подвергаются окислению через цикл ди-трикарбоновых кислот (цикл Кребса). Около 2/3 энергии освобождается в цикле Кребса.
АТФ улавливает и накапливает энергию, освобождающуюся при распаде высокомолекулярных органических соединений в организме. Одновременно в клетке идет синтез АТФ и аккумуляция энергии в ее фосфорных связях. При синтезе белков, а также при функционировании органов и мышц сопряжено идет распад АТФ по месту макроэргических связей с выделением энергии. Образовавшаяся энергия служит источником для синтеза, а также для двигательных процессов.
Из вышесказанного следует, что АТФ является связующим звеном между двумя противоположными процессами, где она при распаде веществ аккумулирует энергию, а при ассимиляции ее отдает.
Биологическую роль АТФ в энергетике обмена можно представить на примере работающего сердца. При взаимодействии с сократительными белками мышц АТФ обеспечивает энергию, необходимую для сокращения сердца и проталкивания крови в кровеносную систему. При этом для бесперебойной работы сердца необходимо постоянное пополнение количества АТФ. Если сердце не получит необходимого количества питательного материала и «горючего» (углеводы и продукты их распада), а также кислорода, необходимого для образования АТФ, то в этом случае наступает нарушение работы сердца.
Необходимое количество АТФ для функционирования различных органов вырабатывается в клеточных организмах – метохондриях в процессе окислительного фосфорилирования.
Практическое занятие № 15.
Задание к занятию № 15.
Тема: ЭНЕРГЕТИЧЕСКИЙ ОБМЕН.
Актуальность темы.
Биологическое окисление – совокупность протекающих в каждой клетке ферментативных процессов, в результате которых молекулы углеводов, жиров и аминокислот расщепляются, в конечном счете, до углекислоты и воды, а освобождающаяся энергия запасается клеткой в виде аденозинтрифосфорной кислоты (АТФ) и затем используется в жизнедеятельности организма (биосинтез молекул, процесс деления клеток, сокращение мышц, активный транспорт, продукция тепла и др.). Врач должен знать о существовании гипоэнергетических состояний, при которых снижается синтез АТФ. При этом страдают все процессы жизнедеятельности, которые протекают с использованием энергии, запасенной в виде макроэргических связей АТФ. Наиболее распространенная причина гипоэнергетических состояний – гипоксия тканей , связанная со снижением концентрации кислорода в воздухе, нарушением работы сердечно-сосудистой и дыхательной систем, анемиями различного происхождения. Кроме того, причиной гипоэнергетических состояний могут быть гиповитаминозы , связанные с нарушением структурного и функционального состояния ферментных систем, участвующих в процессе биологического окисления, а также голодание , которое приводит к отсутствию субстратов тканевого дыхания. Кроме того, в процессе биологического окисления образуются активные формы кислорода, запускающие процессы перекисного окисления липидов биологических мембран. Необходимо знать механизмы защиты организма от данных форм (ферменты, лекарственные препараты, оказывающие мембраностабилизирующее действие – антиоксиданты).
Учебные и воспитательные цели:
Общая цель занятия: привить знания о протекании биологического окисления, в результате которого образуется до 70-8- % энергии в виде АТФ, а также об образовании активных форм кислорода и их повреждающего действия на организм.
Частные цели: уметь определять пероксидазу в хрене, картофеле; активность сукцинатдегидрогеназы мышц.
1. Входной контроль знаний:
1.1. Тесты.
1.2. Устный опрос.
2. Основные вопросы темы:
2.1. Понятие об обмене веществ. Анаболические и катаболические процессы и их взаимосвязь.
2.2. Макроэргические соединения. АТФ – универсальный аккумулятор и источник энергии в организме. Цикл АТФ-АДФ. Энергетический заряд клетки.
2.3. Этапы обмена веществ. Биологическое окисление (тканевое дыхание). Особенности биологического окисления.
2.4. Первичные акцепторы протонов водорода и электронов.
2.5. Организация дыхательной цепи. Переносчики в дыхательной цепи (ЦПЭ).
2.6. Окислительное фосфорилирование АДФ. Механизм сопряжения окисления и фосфорилирования. Коэффициент окислительного фосфорилирования (Р/О).
2.7. Дыхательный контроль. Разобщение дыхания (окисления) и фосфорилирования (свободное окисление).
2.8. Образование токсичных форм кислорода в ЦПЭ и обезвреживание перекиси водорода ферментом пероксидазой.
Лабораторно-практические работы.
3.1. Методика определения пероксидазы в хрене.
3.2. Методика определения пероксидазы в картофеле.
3.3. Определение активности сукцинатдегидрогеназы мышц и конкурентное торможение её активности.
Выходной контроль.
4.1. Тесты.
4.2. Ситуационные задачи.
5. Литература:
5.1. Материалы лекций.
5.2. Николаев А.Я. Биологическая химия.-М.: Высшая школа, 1989., С 199-212, 223-228.
5.3. Березов Т.Т., Коровкин Б.Ф. Биологическая химия. - М.: Медицина, 1990.С.224-225.
5.4. Кушманова О.Д., Ивченко Г.М. Руководство к практическим занятиям по биохимии.- М.: Медицина, 1983, раб. 38.
2. Основные вопросы темы.
2.1. Понятие об обмене веществ. Анаболические и катаболические процессы и их взаимосвязь .
Живые организмы находятся в постоянной и неразрывной связи с окружающей средой.
Эта связь осуществляется в процессе обмена веществ.
Обмен веществ (метаболизм)– совокупность всех реакций в организме.
Промежуточный обмен (внутриклеточный метаболизм) – включает 2 типа реакций: катаболизм и анаболизм.
Катаболизм – процесс расщепления органических веществ до конечных продуктов (СО 2 , Н 2 О и мочевины). В этот процесс включаются метаболиты, образующиеся как при пищеварении, так и при распаде структурно-функциональных компонентов клеток.
Процессы катаболизма в клетках организма сопровождаются потреблением кислорода, который необходим для реакций окисления. В результате реакций катаболизма происходит выделение энергии (экзергонические реакции), которая необходима организму для его жизнедеятельности.
Анаболизм – синтез сложных веществ из простых. В анаболических процессах используется энергия, освобождающаяся при катаболизме (эндергонические реакции).
Источниками энергии для организма являются белки, жиры и углеводы. Энергия, заключенная в химических связях этих соединений, в процессе фотосинтеза трансформировалась из солнечной энергии.
Макроэргические соединения. АТФ – универсальный аккумулятор и источник энергии в организме. Цикл АТФ-АДФ. Энергетический заряд клетки.
АТФ – является макроэргическим соединением, содержащим макроэргические связи; при гидролизе концевой фосфатной связи выделяется около 20 кдж/моль энергии.
К макроэргическим соединениям относятся ГТФ, ЦТФ, УТФ, креатинфосфат, карбамоилфосфат и др. Они используются в организме для синтеза АТФ. Например, ГТФ + АДФ à ГДФ + АТФ
Этот процесс называется субстратное фосфорилирование – экзоргонические реакции. В свою очередь все эти макроэргические соединения образуются при использовании свободной энергии концевой фосфатной группы АТФ. Наконец, энергия АТФ используется для совершения различных видов работ в организме:
Механической (мышечное сокращение);
Электрической (проведение нервного импульса);
Химической (синтез веществ);
Осмотической (активный транспорт веществ через мембрану) – эндергонические реакции.
Таким образом, АТФ- главный, непосредственно используемый донор энергии в организме. АТФ занимает центральное место между эндергоническими и экзергоническими реакциями.
В организме человека образуется количество АТФ, равное массе тела и за каждые 24 часа вся эта энергия разрушается. 1 молекула АТФ «живет» в клетке около минуты.
Использование АТФ как источника энергии возможно только при условии непрерывного синтеза АТФ из АДФ за счет энергии окисления органических соединений. Цикл АТФ-АДФ – основной механизм обмена энергии в биологических системах, а АТФ – универсальная «энергетическая валюта».
Каждая клетка обладает электрическим зарядом, который равен
[АТФ] + ½[АДФ]
[АТФ] + [АДФ] + [АМФ]
Если заряд клетки равен 0,8-0,9, то в клетке весь адениловый фонд представлен в виде АТФ (клетка насыщена энергией и процесс синтеза АТФ не происходит).
По мере использования энергии, АТФ превращается в АДФ, заряд клетки становится равным 0, автоматически начинается синтез АТФ.